Some thoughts on how we can transition from the simple, cell-to-cell editor to dealing with a macroscopic body plan. Parts of this concept depend on talk related to how the editor will work, so it isn’t a self-contained concept. But it should serve what I think is a robust outline for how we can transition out of the microscope.
THE TRANSITION TO COMPLEX MULTICELLULARITY
As Nunz stated, we are on the backend of development for the Microbe Stage. We are focusing less and less on implementing small pieces and are focusing more and more on the puzzle as a whole. As such, it is important to identify the general idea for transitioning away from the Microbe Stage and towards the Multicellular Stage. That way, as we start filing down the rough edges of the current stage, we can be assured that we are heading towards the same direction, and that that direction is the best possible way forward.
The biggest challenge represented by the Multicellular Stage is the transition from unicellular/simple multicellular editing of individual cells to dealing with an entire organism. We obviously can’t charge the player with having to finetune every single cell type that goes in a complex organism’s morphology; that would require a lot of individual, incredibly in-depth parts, such as contractile vacuoles and neuron sheaths, digestive cilia, etc., and would require a lot of knowledge on the hand of the player to adequately put together a creature out of these endless cellular variations. There are pretty long-established concepts regarding this need to simplify things - namely, that the editor will transition away from editing individual cells and instead editing tissues. But beyond that brief synopsis, there is barely any meat to the concept (as far as I am aware). But have no fear, for Deus has a scheme.
My idea essentially would involve incorporating germ layers into the game. Germ layers serve as the backbone of embryology, but they are incredibly important in phylogenetic analysis of Metazoans as a whole, and have very profound insights on evolution.
I’d like to note two things. First, while I am decently familiar with germ layers as a whole, embryology and developmental biology as a whole are incredibly complex and difficult topics, so I am by no means very well-versed in the topic and would really appreciate input from theorists and the team as a whole. And second, while I believe plants and fungi have analogous cell differentiation structures, I haven’t looked into them yet. So this addresses the animal-analogue part of Thrive. I plan to address plant evolution when I add on to the sessile gameplay concept I shared with you guys on Discord.
BACKGROUND
Germ layers are most commonly used in the context of embryo development. They refer to one of three layers of cells in an embryo, and are important because each of these layers specializes into different types of cells.
There are three germ layers: the endoderm, the mesoderm, and the ectoderm. The endoderm is the innermost layer of cells. The cells which form the endoderm differentiate into the digestive tract (including the mouth opening and the anus opening) and digestive organs, the basis of the respiratory tube, the endocrine system, auditory system, and urinary system. The mesoderm is the middle layer of cells, and mesodermal cells differentiate into muscles, bones, the circulatory system, and various other advanced tissues that augment other organ systems, such as developing into lung/gill muscles from the endodermal root provided by the respiratory system. And the ectoderm, the outermost layer, differentiates into cells which form the epidermis and the nervous system (including the brain and spinal cord).
The Embryo Project Encyclopedia is a great source of information for the above concepts. Here is a page for the Endoderm, and other pages can be accessed through the search bar: Endoderm | The Embryo Project Encyclopedia
Germ layers are evolutionary significant because they are important phylogenetic markers. When we talk about organisms possessing differentiated tissues, we are really talking about organisms possessing germ layers. Sponges technically have at most 12 different “types” of cells, but aren’t considered to have differentiated tissues because they don’t have germ layers - actually, they technically only have a single germ layer, but it can’t really be classified as endodermal, mesodermal, or ectodermal in the same way more complex body plans can be. The most simple animals - Porifera, the phylum which encompasses the sponges - only have a single germ layer. Ctenaphora and Cnidaria contain two germ layers, the endoderm and the ectoderm, and are referred to as diploblastic animals. All other animal groups - the remaining invertebrates and the vertebrates - are triploblastic, having the ability to develop muscles and skeletons.
An area of research involves tracing the evolution of endoderm, mesoderm, and ectoderm throughout phylogeny. Diploblastic organisms evolved from multicellular animals without dedicated germ layers, and triploblastic organisms evolved from diploblastic animals - that much is clear. There is a loose understanding of how diploblasty evolved, and I can pull up more research for that later. The gap between diploblasty and triploblasty, however, is a bit less easy to span because modern diploblastic and triploblastic organisms appear to be very different. It is likely that a flatworm-like creature was the basal triploblast. Platyhelminthes are considered to be the most simple of all triploblasts, lacking a dedicated circulatory or respiratory system.
HOW IT CAN WORK
At some point in the early multicellular stage, the player reaches a threshold; instead of individually dealing with editing types of cells and placing each individual cell down in the editor, a general body-plan is presented (what is this threshold; number of cells, an unlock?).
In their next trip to the editor, instead of the familiar microscopic interface, the player is looking at a very simple, soft-bodied wormlike blob. Although their previous cellular composition influences the abilities this worm creature has - if they had toxins they retain that ability to inject/secrete, if they had mitochondria they are aerobic, etc. - they are no longer able to place organelles/proteins. Instead, the player notices they are now able to interact with larger parts - appendages, eyespots, and more (we need to think up of some of the most basal components we’ll offer the player in the earliest macroscopic stages). They’ll also notice the editor has changed slightly: the three tabs are now “Structure”, containing the parts to be placed, “Body Plan”, which I will describe, and “Behavior”, which affects the organism’s behavior.
“Body Plan” replaces the “Membrane” tab, and it deals with widespread changes to the germ layers your creature has rather than placing specific parts. Because the basic macroscopic organism is diploblastic, that would mean there would be two “sections”: the Ectoderm and the Endoderm.
The Ectoderm deals with the “skin” of your organism, so it behaves somewhat similarly to the previous Membrane tab. Just as membrane types previously existed in the microbe/early-multicellular stages, similar membrane types also exist in the macroscopic editor. For example, a chitinous ectoderm would be analogous to flexible exo-skeleton as is seen in arthropods, a calcium carbonate ectoderm would indicate the creation of a rigid yet robust shell as is seen in clams and snails, and double ectoderm would be analogous to “basic”, uncovered yet versatile skin as is seen in most animals. Ectodermal thickness can have implications on movement and temperature resistance, and
The endoderm will represent the start of messing with the organ systems with your organism, and contains the roots of various important systems within them. Containing the roots of the digestive tract, the respiratory system, (and the endocrine system if we see the need to include it/come up with fun gameplay options), some limited customization options will be present already in diploblastic organisms. For example, the digestive tract can allow customization of diet based on membrane, and can be customized to allow more storage and somehow correspond to digestive efficiency (how quickly you extract energy from food in your gut). Perhaps more storage means less efficiency while less storage means more efficiency? There can be many ways of dealing with this.
The basal respiratory system can have a slider related to oxygen-intake efficiency. Higher oxygen-intake efficiency gives you more energy quickly but burns through your energy source quicker, meaning you’ll need to be more ravenous, and also meaning your organism doesn’t do well in low-oxygen settings, such as the deeper ocean. Slower oxygen-intake efficiency gives you less energy but burns through your energy source slower, meaning you’ll have slower metabolism while also perhaps allowing you to live in the ocean depths. There is a bunch of customization options with the respiratory system I can envision related to invertebrate respiratory systems, but let’s make sure the basic idea of this concept is accepted before we spend a lot of time on that.
The mesoderm will of course be missing at first in diploblastic organisms. Until then, the area in between the endoderm and ectoderm will be filled by mesoglea - the gelatinous material found in Cnidaria and Porifera which can be composed of both acellular and cellular material (question for theorists: is mesoglea assumed to be basal for all diploblastic animals or is it a unique adaptation to certain diploblasts?). A lack of mesoderm means the player will have limited options in regards to functionality drawn from mesoglea. However, one important function can be related to the organism’s density, and thus, whether your diploblastic animal drifts or crawls on the ocean floor: the more mesoglea, the less energy your organism needs to float above the surface of the water. As such, we can have a slider denoting the amount of mesoglea in your organism for now - more mesoglea means a lighter, more floaty organism akin to jellyfish, more agile and nimble yet more vulnerable to currents and with less health, while less mesoglea means a denser, more durable benthic organism with limited ability to get off the ocean floor. When the player evolves mesodermal cells, the mesoglea’s functions (density) will be replaced by some of the added customization options - for example, bone density and air bladders composed of mesodermal cells can fill in mesoglea’s function.
Adopting the mesoderm will replace this mesoglea with muscles, and will almost serve as the “nucleus” of the late multicellular stage, catapulting the player towards the early Aware stage. The mesoderm will allow the roots of organ systems provided by the endoderm to expand in creativity and function, increasing the efficiency of the respiratory system with an upgrade and allowing the adaptation of advanced gills and lungs. The evolution of mesoderm will allow the evolution of bone tissue, allowing the evolution of a spinal cord, and eventually, parts like vertebrate limbs, skulls, teeth, etc.; and for organisms with chitin, the evolution of advanced muscles will allow the evolution of segmentation and arthropod appendages. There’s a lot we can discuss with the mesoderm, but again, for now, I want to make sure the basic idea makes sense.
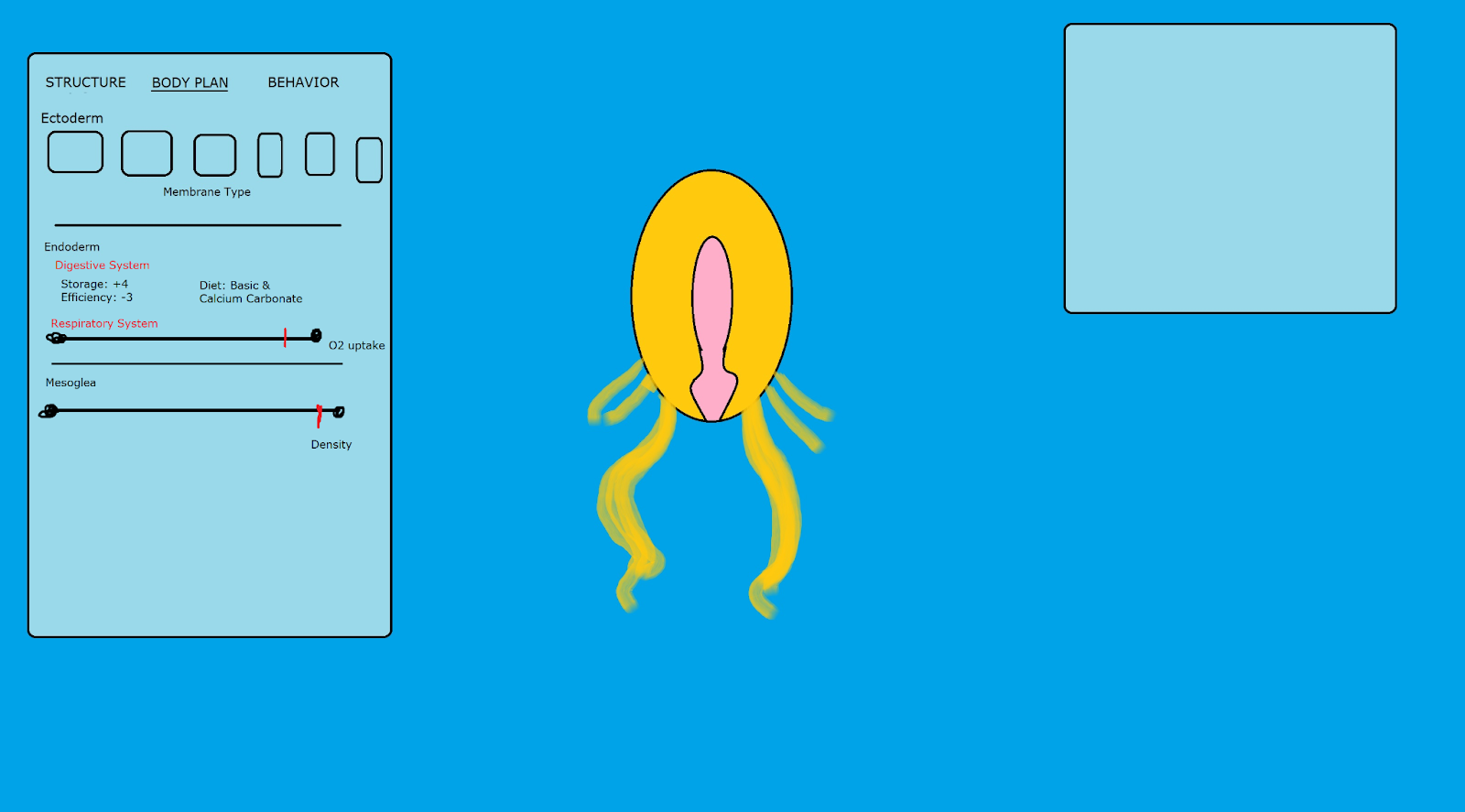
A very basic and sucky Paint concept from me of how it can look like with a ctenaphore-inspired analogue. I tried to create a GUI for the editor, but I’m not very good as you can see.
Note: I didn’t address mesoderm a lot for a reason. The mesoderm really allows the proliferation of organ systems as you might see from me mentioning bones, circulatory system, etc. Many organ systems involve a root provided by the endoderm and expanded upon by mesodermal tissue. There’s a lot of flexibility there of course, but I want to make sure the basic idea is robust enough for you guys, and I want to make sure that the closer future of Thrive is more perfectly addressed.
QUESTIONS AND IMPLICATIONS
I think this idea presents a very flexible and game-friendly way to represent the evolution of an organism from a simple multicellular organism to a complex organism with a general body plan. However, there are still questions remaining. This concept of course relates heavily with concepts for the 3D editor, so there’s a bit of a gray area where I’m not exactly sure how to deal with specific cool adaptations we see in life today.
Here are a list of questions and observations…
- Sponges and the Such - Sponges only have a single germ layer, but are still macroscopic organisms. This ties into the conversation surrounding sessile gameplay, and I had some thoughts about that I shared with you guys, but how could we represent such organisms?
- Ectoderm Details - How do we deal with, and balance, the player having multiple types of ectoderm? For example, a player who wants a shell they can retract into would probably be eying calcium carbonate or chitin, but they wouldn’t want all of their organism’s ectoderm to be rigid; they’d still want a soft-bodied part. Perhaps a specific part in the Structure tab can deal with this?
- Transitions, Transitions, Transitions - How will we incorporate the transition to diploblasty, and from diploblasty to triploblasty? For the former, I’d assume we’d want something else rather than “get x amount of cells and click a button in gameplay”. For the latter, it’s a matter of limiting progression fairly. I didn’t cover the nervous system here although it is a component of the ectoderm; perhaps developing a very simple nervous system will transition you to editing a body plan, and upgrading the nervous system will transition you to triploblasty? But then there’s the question of how to represent the nervous system; maybe it can be an upgrade in the Body Plan section of the simple multicellular stage as is seen in game now, where you can click a button that automatically creates a nervous system after you are generating a certain amount of ATP?
- Plants and Animals - Plants have analogous structure to germ layers within them: dermal, ground, and vascular tissue. How do we deal with that transition for that path of life? Perhaps we should rename “germ layers” to something a bit more neutral, and have different adaptations for organisms that have chloroplasts. Ties into the question of sessile gameplay/editors.
- Different “Types” of Editors - I put the word types in quotation marks because we just need different “tools” within the same macroscopic editor instead of completely different editors. I think we’d want different “tools” for soft-bodied organisms like jellyfish and worms and molluscs and the such, vertebrate organisms with internal skeletal structures, and exo-skeleton organisms with external skeletal structures such as in arthropods. For example, vertebrates would inherently edit their morphology first by adding bones for limbs and skulls and the such; it wouldn’t be realistic to have a arthropod-analogue organism be edited through similar tools because arthropods edit morphology through segmentation rather than through internal skeletal editing. The good news is that there are analogues and overlaps. For example, nothing stops a soft-bodied appendage, such as gills on axolotl, from appearing on vertebrates, limbs across vertebrates and arthropods are basically identical, etc.
- Worm World - I think the basic “template” organism we can expect to see in Thrive is a worm-like organism; for example, I anticipate the player’s basic animal which emerges from the microscopic-macroscopic editor transition to be a worm-like creature. From this worm, different customization options can be attached to allow the player and auto-evo to expand in morphology. A somewhat similar trend is apparent in real evolutionary history - there appears to have been a “worm world” a bit before the Cambrian.
- Organ System Variance - There are differences in organ systems observed across Eumetazoans. For example, arthropods have an open circulatory system whereas most other “higher” animals have a closed, tubular circulatory system. An open digestive system has an entrance (the mouth) and an end (the anus) whereas a closed digestive system only has one opening for both waste and input. We can have “variants” of organ systems with their own attached benefits and shortcomings, so that could be a future area of much discussion.
One last note: I think a good way to measure the strength of a concept related to the editor/progression question is asking yourself if the concept can realistically create a specific unique organism you have in mind. For example, if an editor concept can create a coral, a jellyfish, an insect, and a vertebrate, I feel like that’s a robust concept.