I realized I made a mistake with the corrected diagram at the top of this post. It should actually look like this:
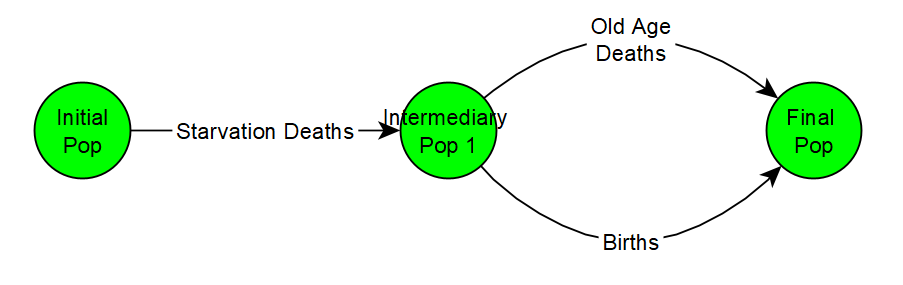
In the updated algorithm, the first thing that happens is that the script determines what fraction of the population dies from starvation and kills them off first. Then of the remaining population, it determines how many offspring were birthed. Before it adds the births, it also kills off any of the population members who die from old age. Therefore births and old age deaths occur sort of in parallel.
Anyways, just wanted to make that clear and I’ll go back and fix the original post later. The next part will be coming up soon! Feel free to leave any suggestions or feedback you have on the series so far.