As has come up recently in Multicellular design discussions, we will probably end up having a cell adjacency system. Since that will be implemented in the base hex editor, it will automatically also be available for the cell editor. Which means we can either have mechanics for adjacency between cell parts at a relatively low cost, or deliberately disable it. We did at least have some prior community requests for more depth in the editor experience, though it has been a while I believe.
Here, I’ve written up a design concept as good as I could think of, so that we can decide if it would be a nett positive to the game experience.
Theoretical background
First, since this is Thrive, let’s look at the real life patterns.I looked at a variety of diagrams (to see how the field perceives what cells look like) and electron micrographs (to make sure that also matches reality). Some examples here:







To, me most organelles look quite evenly distributed. There are some “required” adjacencies/proximities, like the nucleus + ER (endoplasmatic reticulum) + Golgi (all one part in Thrive). But most can be found roughly “everywhere”. In cases where they’re not equally distributed it seems like that’s either because there’s literally only one in the cell, or because they are needed in a certain location of the cell relative to the outside world. Organelles of the same type don’t cluster together by default.
Side note on prokaryote parts.
Note that here I am talking about relatively large membrane-contained organelles. (mainly eukaryotic parts in Thrive). For much smaller components and simple proteins (most of the early parts in Thrive), you’re just not going to get as clear a picture:
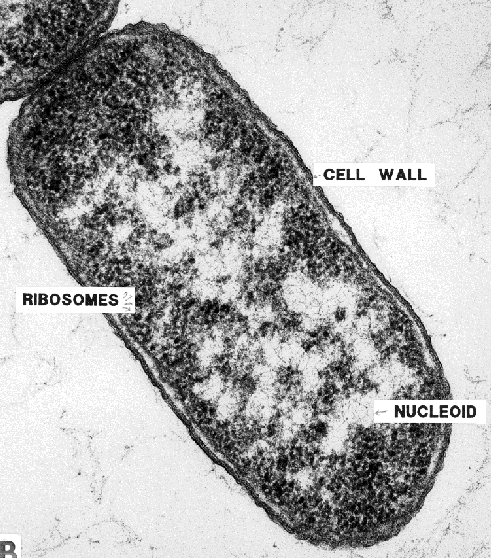

With perhaps some exceptions, like:


It would be easy to think that those organelles randomly float around. But we do know that is not true and organelles are very “deliberately” positioned in the cell, mainly through action of the cytoskeletal system.

And moving organelles around is used quite a lot, for example concentrating mitchondria when lots of ATP is required on one side of the cell:

So, my conclusion is that the relatively even distribution is deliberate. Chemically, this makes sense. Substance need time to diffuse. Low local concentration of reaction “ingredients” slows down reactions. High local concentrations of products can do the same. The highest concentration of a compound is probably going to be where it is produced.
Then it follows that:
- It is more efficient to have organelles that perform a certain chemical process not all clustered together in one location. (They would be eating each other’s resources)
- It is more efficient to have parts that are consume a compound next to ones that are producing that same compound. (Nearly everything uses ATP, so mitochondria are everywhere.
This does raise some questions. For example, IRL organelles are moved around a lot. So does it make sense to reward players for their exact position? This goes doubly so for the even smaller prokaryotic protein “parts”. Is the scientifically accurate “even distribution” a fun gameplay objective to chase?
A bit of a more unique case is the Nucleus part. If you’ve paid close attention, when looking a cross-sections of a eukaryotic cell, you can see bits of the ER appear away from the Nucleus. It’s still attached in another plane, but it does try to spread out. The nucleus, via de ribosomes in the ER produces the proteins that are the machinery for practically everything that happens in the cell. So you could argue that every cell part should benefit from being adjacent to the nucleus.
Design objectives
A few of the thoughts that came to mind while conceptualising in random order:
- From the theory, what we want to see is a quite even distribution of organelles throughout the cell, but especially a focus on placing production of resources next to consumption of resources.
- Also from the theory, I could imagine a penalty for placing the same organelles next to each other, but that does not seem like a fun experience to me. It would also be harder to adjust to for existing players.
- We would of course prefer to keep the rules simple and consistent for ease of implementation and clarity to the player.
- I think this should be a relatively minor effect, or it would be a lot of pressure on a puzzle game that not everyone might find fun.
- I don’t think it makes sense to involve osmoregulation cost here in any way, since that does not differentiate between organelle types.
The Concept(s)
For both examples below, it would probably be optimal if we can let this depend on the number of adjacent hexes/edges, otherwise some of the optimal positioning of multi-hex organelles next to for example the nucleus could end up feeling artificial.
Nucleus adjacency:
Any part adjacent to the nucleus gets a boost. Speeds up processes, flagella give more speed, vacuoles have more storage, etc. It reminds of some adjacency-based building systems in games that have a “capital building” that gives bonuses to everything directly adjacent. Simple concept, but would need some work to detail and implement bonuses for each organelle. (But perhaps we could use mostly the same bonuses as the cell specialisation system?)
Process adjacency:
For a cell part that turns compound A into compound B:
- Process speed is increased for every adjacent cell part that produces compound A.
- Process speed is increased for every adjacent cell part that consumes compound B.
- Specialised vacuoles could be counted as a cell part that consume and/or produces that compound.
- Every cell part counts as consuming ATP, though most less than those that specifically guzzle ATP, like flagella and bioluminescent vacuoles.
Example “adjacency optimised” cells:


So, that’s what I got so far, I think it’s workable (though I can’t comment on ease of implementation) but I think it should not be tuned overly strong, or it could overshadow other parts of editor gameplay.